
Hering a gândit și a lucrat într-un mod complet diferit de Helmholtz. Helmholtz reușise să ofere o bază fiziologiei vederii cu ajutorul matematicii și fizicii. Hering, mai departe, dimpotrivă, a preferat datele psihologice, încercând să-și imagineze ce procese neuronale erau responsabile de senzații.
Teoria lui Hering se bazează pe o analiză a percepției vizuale. Experiența indică faptul că un subiect conștient are nevoie de patru culori unice pentru a caracteriza percepția: albastru, galben, roșu și verde. De obicei, două dintre aceste nuanțe constituie o senzație de culoare, de exemplu, portocaliul conține roșu și galben, în timp ce albastru și verde sunt componentele cyan-ului.
Ideea centrală din conceptul lui Hering era că roșul și verde sunt nuanțe opuse, deoarece nu sunt niciodată provocate simultan de un stimul de culoare; același lucru este valabil și pentru albastru și galben.
Hering, ca și Goethe, a fost preocupat de unicitatea albului. S-a opus opiniei lui Grassmann (și a lui Fechner) care susțineau că „punctul zero al luminii”. Hering a considerat că negrul este la fel de mult a senzație la fel ca albul. Prin urmare, a dedus că trebuie să existe o substanță vizuală receptivă care să poată suferi o schimbare pozitivă atunci când albul este văzut și o schimbare negativă când se percepe negrul.
El a mai presupus că percepem griurile atunci când acele substanțe sunt în diverse stări de echilibru dintre alb-negru. Practic, Hering a presupus la la nivelul retinei sunt prezente trei perechi de substanțe, pe care el le-e definit ca fiind alb-negru, roșu-verde și albastru-galben. Mai departe, el a sugerat că lumina acționează asupra acestor trei substanțe fotosensibile prin, ceea ce a numit el, asimilare (A) și disimilare (D).
Atunci când lumina acționează într-o manieră de disimilare sau descompunere asupra substanței fotosensibile alb-negru, este produsă sensația de alb, iar atunci când lumina acționează în mod asimilativ sau regenerativ, apare senzația de negru. Hering nu a reușit să determine ce tip de lumină este asimilativă sau disimilativă. Însă, el a presupus că roșul determină un efect de descompunere asupra substanței roșu-verde iar verdele are un efect de regenerare asupra aceleiași substanțe.
Albastrul, considera el, acționează disimulativ asupra substanței albastru-galben, pe când regenerarea e cauzată de culoare galbenă. De asemenea, el a mai presupus, că toate culorile acționează într-un mod disimulativ asupra substanței alb-negru, producând senzația de alb, concomitent culorilor aferente.
Astfel, potrivit teoriei lui Hering, perechile de culori alb-negru, rosu-verde, albastru-galben erau considerate opuse, deoarece o culoare dintr-o pereche o neutralizează pe cealaltă, din aceeași pereche. Relațiile dintre perechile de culori, au fost tratate de Young-Helmhotz ca un proces aditiv, pe când hering le-a tratat din punct de vedere substractiv.
Conform lui Hering, atunci când culoarea roșie și cea verde se amestecă la nivelul retinei, ele se neutralizează reciproc, fiind culori opuse, acționând antagonist. Dar rămâne capacitatea lor de disimilare, care acționează asupra substanței alb-negru, producând senzația de alb. În schimb, teoria Young-Helmholtz, postula că toate culorile, în afară de cele primare, sunt rezultatul amestecului acestora.[1], [2].
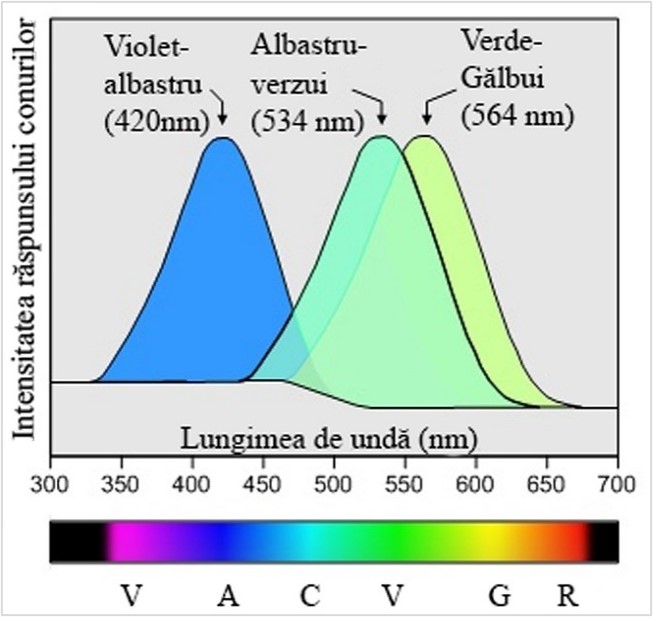
Există aproximativ 6 milioane de celule sensibile la diverse lungimi de undă ale spectrului vizibil în fiecare ochi. Fiecare tip de celulă con acoperă o gamă largă de frecvențe, dar în primul rând ele au o sensibilitate la o anumită porțiune din spectru. Astfel un anumit tip de celulă conică este sensibilă în primul rând la lumina albăstrui-violetă, dar alte două tipuri sunt cele mai sensibile la lungimile de undă corespunzătoare gamei de culoare verde; unul atinge vârfurile la un verde-albăstrui și celălalt la vârfurile la un verde-gălbui.
Fiecare din cele trei tipuri de con (I, II, III) conțin fotopigmenți proteici (-opsine) și sensibilitatea maximă, peak-ul, este dată, separat, de fiecare tip în parte, la lungimi diferite de undă ale spectrului vizibil:[3]
- Undele scurte stimulează conurile S, corespunzătoare spectrului vizibil din regiunea viotet-albastrui (~420nm)
- Undele medii stimulează conurile M, corespunzătoare spectrului vizibil din regiunea verde-albăstrui (~534nm)
- Undele lungi stimulează conurile L, corespunzătoare spectrului vizibil din regiunea verde-gălbui (~564nm)
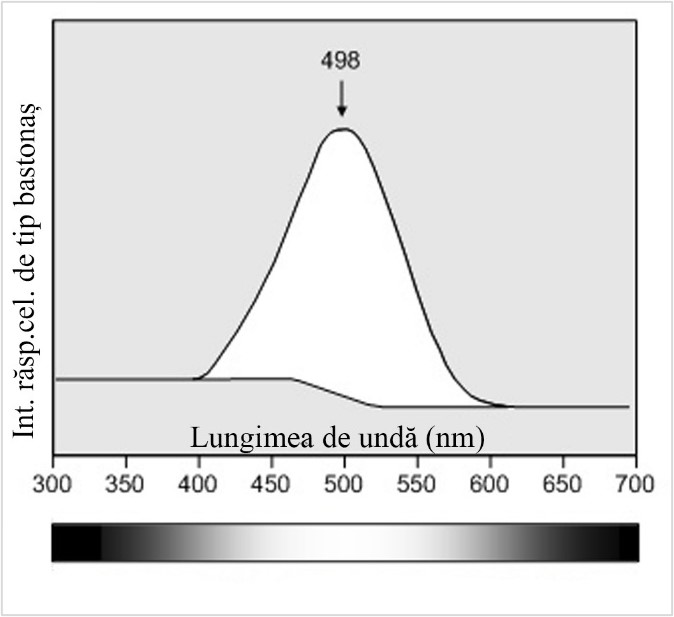
În figura 1 este ilustrat răspunsul conurilor de tip I, II, III (S, M, L) la lungimile de undă ale spectrului vizibil iar în figura 2 este ilustrat, comparativ, răspunsul celulelor de tip bastonaș în condiții slabe de iluminate (vedere nocturnă).
Conurile de tip S reprezintă aproximativ 10% din numărul total de conuri. Celulele bipolare și GC transmit semnalele din conurile S și există dovezi că acestea au o cale de semnal separată prin talamus până la cortexul vizual.
Pe de altă parte, conurile de tip L și M reprezintă restul de 90% din numărul total de conuri. Cu toate acestea, ele sunt greu de distins deoarece au aceeași formă, diferența dintre ele fiind numărul diferit de aminoacizi care formează -opsinele care fac parte din ele.
Mollon și Bowmaker în anul 1992,[4] ulterior Wässle, în anul 1999,[5] au descoperit faptul că distribuția conurilor L și M este aleatorie, în proporție de 1:1.
Teoria culorilor opuse, alternativă, a fost propusă pentru prima dată de fiziologul și psihologul german Karl Ewald Hering (1834-1918).
Inițial teoria nu a beneficiat de o atenție deosebită din partea comunității științifice, dar procesul adversar a recâștigat teren în anii 1970, de alți cercetători.
Sunt două idei care stau la baza procesului adversar, și anume:
- Celulele fotosensibile de la nivelul retinei conțin trei substanțe proteice diferite care se modifică metabolic atunci când fotonii ajung să interacționeze cu acestea, producând următoarele perechi de senzații: alb-negru, rosu-verde și galben-albastru. Informația care ajunge la creier sub formă de impulsuri nervoase codificate, fiind urmare a modificărilor și a interacțiunilor dintre aceste substanțe fotosensibile.
- Sensibilitățile celulelor conice ating peak-ul la frecvențe diferite, există o cantitate mare de suprapuneri în ceea ce privește lungimile de undă ale luminii la care răspund cele trei tipuri de celule conice, astfel încât sistemele noastre vizuale sunt concepute pentru a detecta diferențele între răspunsurile diferitelor conuri la nivelul creierului. Pentru ca aceste diferențe să fie percepute, retina se folosește de un număr mare de celulele ganglionare (GC)[6] – fiecare dintre aceste celule fiind capabile să compare semnalele generate de un număr de celule conice diferite, și semnalele care ies din GC sunt cele care furnizează informații despre culoare creierului. Astfel, percepem culoarea galbenă atunci când conurile sensibile la lungimea de undă corespunzătoare culorii verde-gălbui sunt stimulate puțin mai mult decât conurile sensibile la lungimea de undă corespunzătoare culorii verde-albăstrui, de exemplu; în mod similar, percepem culoarea roșie atunci când conurile sensibile la lungimea de undă corespunzătoare culorii galben-verzui sunt stimulate semnificativ mai mult decât conurile sensibile la lungimea de undă corespunzătoare culorii verde-albăstrui.
Hering a avut dreptate în analiza conform căreia există două perechi de culori de bază care au o relație adversară, unică pentru fiecare pereche. În același timp, ipoteza sa, conform căreia oponența dintre culori se produce la nivelul celulelor fotoreceptoare s-a dovedit a fi incorectă.
Pe vremea lui Hering, înțelegerea neuro-fiziologiei era slabă, iar conceptele de excitație și inhibare a neuronilor în moduri unice și complexe erau necunoscute. Prima sugestie, conform căreia antagonismul dintre culori ar putea fi cauzat de excitația, respectiv inhibarea care are loc la nivelul celulelor nervoase, a fost într-o lucrare a fiziologului suedez Gustaf F. Gothlin (1944).[7]
Acesta a făcut cercetări pe pornind de la concluziile lui Sherington, publicate în 1906 în lucrarea sa The Integrative Action of the Nervous System, care a cercetat procesul de inervație reciprocă și de la care a evoluat ideea de excitație și inhibare între celulele nervoase.[8], [9].
Gothlin sugerează că există într-adevăr un proces antagonist în perechea de senzație albastru-galben ca primă etapă și mai apoi, sugerează că componenta galbenă a balanței albastru-galben este implicată în relația antagonistă dintre roșu și verde. El arată că aceste interacțiuni antagoniste s-au datorat excitației și inhibării dintre celulele fotoreceptoare.
Granit, în 1947, a propus o teorie care implică un complex dominator- modulară a vederii culorilor. Conform teoriei sale, la nivelul retinei există un grup de fotoreceptori care răspund doar unei părți înguste a spectrului vizibil (fotoreceptori modulatori) și un alt grup care răspunde la o bandă largă a spectrului vizibil (fotoreceptori dominatori).[10] El a presupus că celulele modulatoare sunt mai implicate în vederea culorilor, dar nu a menționat niciodată conceptul de excitație și inhibiție neuronală la nivelul conurilor, așa cum sugerase Gothlin.
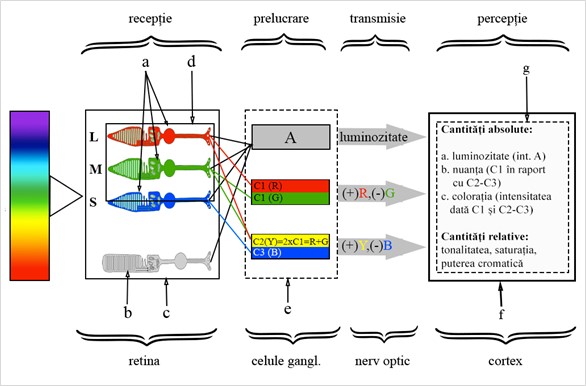
Hurvich și Jameson în 1957, au propus că cele trei tipuri de receptori de culoare – roșu, verde și albastru sensibile – sunt interconectate într-un proces opus la nivelul GC.
Astfel, radiația electromagnetică cu lungime de undă lungă excită conurile „roșii” și celulele ganglionare responsabile de prelucrarea, codificarea fluxului luminos adiacent, roșu-verde – C1(R). pentru a transmite mai departe informația, prin nervul optic, către cortexul vizual unde va forma percepția roșului. Lumina cu lungime de undă medie excită conurile „verzi” și inhibă până la jumătate aceeași celulă ganglionară, reducând excitația acesteia pentru culoarea roșie, reușind astfel să producă percepția luminii verzi – C1(G). La fel, lumina cu lungime de undă scurtă excită conurile „albastre” și inhibă parțial celula ganglionară Y(yellow)B(blue) – C3B, conducând la o senzație de albastru. Lumina, la jumătatea distanței dintre sensibilitățile conurilor „roșii” și „verzi” (R,G) le-ar stimula pe amândouă. Rata de transmisie a impulsurilor în celula ganglionară R(red)G(green) nu s-ar schimba, deoarece stimularea și excitația egală din cele două conuri s-ar anula. Însă conexiunile acestor conuri la celula ganglionară YB duc la excitația acesteia – C2(Y).[11] Astfel, excitația combinată, dată de conurile R și G, produce senzația colorii galbene. Acest proces este ilustrat în figura 3.
Potrivirile de culoare ale cantității Cs ale unui stimul S pot fi exprimate ca:
Cs (S) = C1 (l1) + C2 (l2) + C3 (l3)
unde li reprezintă cele trei culori primare (nu neapărat culori spectrale monocromatice) și Ci sunt valori tristimulus.
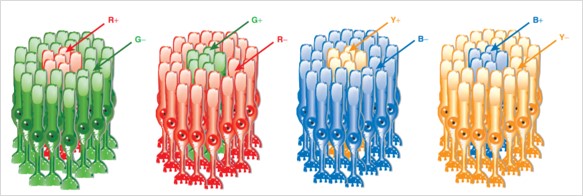
Primele studii care au vizat răspunsurile izolate la stimuli monocromatici ale unui singur fotoreceptor, au fost obținute prin experimente la nivelul nucleului geniculat lateral (LGN) la maimuțe, de către Russel DeValois în 1958.[12]
Acesta a demonstrat prezența celulelor care răspund la un spectru îngust al radiației electromagnetice vizibile bandă îngustă, care corespund celulelor modulatoare descrise de Granit. Totodată, DeValois a arătat prezența unor celule fotoreceptoare care răspund la radiația vizibilă corespunzătoare culorii roșii dar care nu sunt excitate de radiația vizibilă corespunzătoare culorii verzi, indicând astfel o relație antagonistă dintre roșu și verde (fig 4).
Experimentele care au readus teoria lui Hering în atenția cercetătorilor, au fost realizate de Gunter Svaetichin, în 1956, care a folosit sticlă în loc de electrozi metalici pentru a înregistra activitatea intracelulară a conurilor. Astfel, Svaetichin a observat o secvență de răspunsuri depolarizante și hiperpolarizante, corespunzătoare excitației și inhibiției neuronale.[13]
Toate aceste teorii explică vederea tricromată și converg, din punct de vedere științific,[14] explicând în ce mod sunt excitate conurile fotosensibile S, M, L și cum se generează impulsurile nervoase spre creier, locul unde variațiile diferite ale intensității nivelului de excitare mediate de GC comparatoare, fac posibilă percepția tuturor nuanțelor – atât ca și saturație și intensitate – pe care le poate forma amestecul celor trei culorilor fundamentale.
Bibliografie:
[1] Swan M. Burnett, „Color-Blindness and color-perception” în Popular Science, Editura: Bonnier Corporation, 1882 vol. 21, p. 90.
[2] Deane Brewster Judd, Contributions to Color Science, Ed. University of California, 1979, pp. 585-586.
[3] Schnapf JL, Kraft TW, Baylor DA. „Spectral sensitivity of human cone photoreceptors” în Nature., 1987 Jan 29-Feb 4;325(6103):439-41. doi: 10.1038/325439a0. PMID: 3808045.
[4] Mollon, J. D.; Bowmaker, J. K., „The spatial arrangement of cones in the primate fovea” în Nature, 1992, nr. 360 (6405), pp. 677–679.
[5] Wässle, Heinz, „Colour vision: A patchwork of cones”. Nature, 1999, nr. 397 (6719), pp. 473–47.
[6] Notă: Liviu Lăzărescu, în Culoarea în artă, la pagina 32, descriind teoria culorilor opuse, le descrie ca fiind un „al patrulea tip de con, receptiv la variațiile de luminozitate,” în timp ce Wagner, Griggs, Schmidt ș.a. numesc responsabile de variațiile de luminozitate „celule ganglionare” (vezi: Wagner, 1960:1314; Griggs, 2009:92; Schmidt et al., 2014:1-16)
[7] Gustaf F. Göthlin, „Experimental Determination of the Short Wave Fundamental Color in Man’s Color Sense” în Journal of the Optical Society of America, 1944, Vol. 34, Nr. 3, pp. 147-158
[8] Breathnach CS. Charles Scott Sherrington’s Integrative action: a centenary notice, în Journal of the Royal Society of Medicine, 2004, 97(1):34-6.
[9] Finger S. Minds Behind the Brain. New York, NY: Oxford University Press; 2000.
[10] Pickford R. W., Human Colour Vision and Granit’s Theory în Nature, 1948, nr. 162, pp. 414–415.
[11] Hurvich, L.M., Jameson, D. „Color theory and abnormal red-green vision.” în Doc Ophthalmol 16, 409–442 (1962).
[12] Russell L. De Valois, Karen K. De Valois, „A multi-stage color model” în Vision Research, Vol. 33, Nr. 8, 1993, pp. 1053-1065, ISSN 0042-6989. (https://www.sciencedirect.com/science/article/pii/004269899390240W).
[13] Svaetichin, G. and MacNichol, E.F., Jr., „RETINAL MECHANISMS FOR CHROMATIC AND ACHROMATIC VISION”, în Annals of the New York Academy of Sciences, 1958, Nr. 74: 385-404.
[14] Turner, R. Steven. “Vision Studies in Germany: Helmholtz versus Hering.” în Osiris, vol. 8, 1993, pp. 80–103. JSTOR, www.jstor.org/stable/301696. Accesat 17 Mar. 2021.


